There are several strains of Fusarium capable of producing the mycotoxin deoxynivalenol (DON) or vomitoxin, a type-B trichothecene. It should be noted that the common active group of all trichothecenes in the epoxide are responsible for the binding of DON to ribosomes that inhibit protein synthesis.
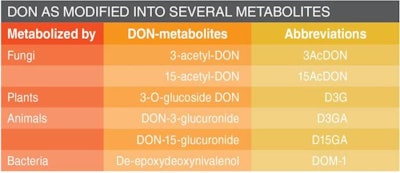
In contrast, the non-toxic de-epoxidated metabolite DOM-1 cannot bind to ribosomes due to the lacking epoxide group. It also is important to know that DON can be further modified into several different metabolites by fungi, plants, animals and bacteria (see Table 1). These DON derivatives also are called masked mycotoxins.
Studies have shown that the amount of DON derivatives, mainly 3/15AcDON and D3G, can account for an additional up to 75 percent of DON contamination in pig feed. Newly released wheat cultivars, which are able to more efficiently convert DON to D3G, are more resistant towards the DON producing fungus Fusarium graminearum, but can contain up to 10-times more D3G than DON.
So, what happens to DON and its derivatives when they enter the gastrointestinal tract of a pig?
DON after ingestion
The intestinal absorption of DON and its metabolites differ between animals. About 54-89 percent of DON can cross the intestinal epithelium and are detected in the blood. The localization of the gut microbiota before the small intestine has a major effect on bioavailability, as DON is mainly absorbed in the small intestine.
In pigs, one of the animals most sensitive to DON, the microbial biomass in the stomach, which is located before the small intestine is in the range of 102-103 per mL intestinal fluid, as shown in Figure 1.
Intestinal bacteria can transform the DON derivatives D3G, 3/15AcDON into DON. But, the transformation of DON into non-toxic DOM-1 by certain bacteria prevents the absorption of DON, and such bacteria are currently commercially available for in-feed use.
After ingestion of DON-contaminated feed, the intestinal epithelial cells are the first target of DON. Regardless of the amount of DON being absorbed, the intestinal epithelium is exposed to the entire contamination of the feed and therefore non-absorbed toxins can also compromise the entire intestine. Absorbed mycotoxins can re-enter the intestine through the intestinal epithelium or through enterohepatic circulation (excretion via bile and re-absorption), thereby increasing the exposure time along the gastrointestinal tract.
DON impairs intestinal barrier functions
The intestinal tract represents an important barrier to ingested chemicals, feed contaminants and is the first line of defense against intestinal infection. The gut barrier is formed to a large extent by tight junctions that seal the luminal end of the intercellular space, as shown in Figure 2.
DON crosses the intestinal mucosa paracellularly through the tight junctions. At the same time, DON increases the paracellular permeability of the intestine through the opening of the tight junctions. Therefore, chronically exposed animals have a higher DON uptake.
This damage in gut health can cause bacteria to be translocated across the intestinal epithelium, increasing the risk of intestinal bacterial infections. In addition, other mycotoxins, pharmaceuticals, pesticides, allergens, fungi and viruses also are granted an easier passage across the intestinal epithelium.
Poor intestinal function reduces nutrient uptake
Seventy percent of the immune system is located in the gastrointestinal tract. DON harms the innate immunity by:
- Direct activation of signal pathways
- Opens tight junctions allowing luminal bacterial antigens to trespass
- Reduces mucus production
High doses of DON repress the immune response, whereas low concentrations promote a rapid mucosal inflammatory response. This poses the risk of induced chronic intestinal inflammation, such as inflammatory bowel disease.
DON also interferes with the intestinal absorption of nutrients, like glucose and amino acids. The sodium-glucose dependent transporter (SGLT-1) is responsible for glucose uptake. Low concentrations of DON are enough to inhibit SGLT-1 and reduce glucose uptake. SGLT-1 is the most DON-sensitive transporter, followed by GLUT-5, the passive fructose transporter.
SGLT-1 is also responsible for water reabsorption and the blocking of SGLT-1 by DON could be the mechanism behind the oft-occurring, DON-induced diarrhea. Low doses of DON reduce the height of the intestinal villi, causing villus fusion and atrophy in the duodenum and jejunum of pigs. Villi increase the internal surface area of the intestinal wall and are therefore necessary for effective nutrient absorption.
DON symptoms: feed refusal, anorexia
Two well-known effects of DON in pigs are anorexia and feed refusal. The mechanisms behind these effects are complex. Scientific evidence shows that within the gut-brain axis, neuroendocrine factors, pro-inflammatory cytokines and bitter taste receptors found throughout the gastrointestinal tract are involved in DON-induced feed refusal. The brain, particularly the hindbrain including the area postrema and hypothalamus, can signal immediate changes in food intake.
As DON is able to cross the blood-brain barrier, about 25-30 percent of the plasmatic DON can be found in the cerebro-spinal fluid of pigs after 2-60 min. On the other hand, DOM-1 cannot cross the blood-brain barrier. A recent study revealed that DON can alter brain functions and directly target the brain causing vomiting, anorexia, fever, decreased locomotor activity and social withdrawal.
Pigs and DON
Pigs are highly sensitive to DON. About 54-89 percent of DON can cross the intestinal epithelium in pigs and are detected in the blood. As intestinal cells are the first cells to be exposed to DON and at much higher concentrations than other tissues, it is of significant interest to investigate how the gastrointestinal tract is compromised by feed contaminated with DON. Finally, it should be kept in mind that the effect on the intestine is only one part of the consequences in animals fed DON-contaminated feed.















