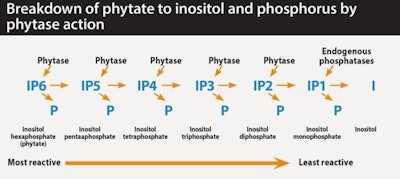
generation E.coli phytases are able to target 90 percent destruction of both phytate (IP6 – phytate is inositol hexaphosphate) and the intermediate lower esters (such as IP4 and IP3) produced as phytate is stripped of P by phytase action, and which also have anti-nutrient effects.
Interest in the benefits of phytase application beyond the simple release of phosphorus (P) from plant phytates is growing rapidly. The use of higher phytase doses to eliminate the anti-nutrient effects of phytate and substantially improve bird performance – superdosing – has expanded to account for 7 percent of the phytase market by value since the concept was launched in 2012.
Three of the top five global monogastric feed producers are now adopting the technique, and within the Brazilian broiler sector, for example, where penetration is highest, more than 20 percent of all phytase-treated feed is superdosed.
Performance benefits
However, the latest research suggests that the performance benefits achieved from superdosing are due to more than simply the elimination of phytate anti-nutrient effects. This discovery is extremely important, and has implications for successfully selecting a phytase suitable for superdosing, and subsequently implementing a superdosing strategy.
Understanding the various mechanisms involved in delivering a superdosing response is therefore vital if the typical three- to four-point improvement in broiler feed conversion ratio (FCR) – worth around US$7-8 per ton of feed – is to be reliably achieved. The same is true for the corresponding response seen in layers, where increased yolk mineral content, increased shell quality, and reduced hen mortality in the later stages of lay represent valuable gains.
Phytate destruction
The first step is to accurately define superdosing itself. For poultry, it is the application at high doses (typically three to four times the standard dose) of an intrinsically thermostable, highly efficient phytase developed specifically to target near-complete phytate destruction. Critically, only a standard-dose mineral matrix is applied during diet formulation.
New generation E.coli phytases are able to target up to 90 percent destruction of both phytate (IP6 – phytate is inositol hexaphosphate) and the intermediate lower esters produced as P is stripped from phytate by phytase action, such as IP4 and IP3. These lower esters also act as anti-nutrients and together, with phytate, the effects are numerous.
For example, phytate will bind to positively charged minerals and feed proteins within the gut and so make them less available. It has also been shown to reduce the activation of the stomach enzyme pepsin, which is responsible for protein digestion.
The resulting rise in undigested protein reaching the small intestine then increases endogenous losses by triggering production of additional hydrochloric acid and pepsinogen (the precursor to pepsin) in the stomach and proventriculus. Further losses come from greater secretion of mucus (to protect the gut wall from the irritant effect of this acid), and sodium bicarbonate (to neutralise the extra acid).
There is also evidence to suggest that phytate negatively affects the mechanism by which amino acids are absorbed from the small intestine, while IP3 and IP4 have been shown to interfere with pepsin activity and reduce the uptake of zinc, calcium and copper.
Inositol importance
These additional anti-nutrient effects highlight how essential it is to achieve phytate degradation below IP3 when superdosing. Just as importantly, it is now becoming apparent that up to 30 percent of the superdosing response seen in broilers may be due to the beneficial effects of the inositol produced after the removal of P from phytate by phytase action.
The growth promoting effects of inositol in chicks have been recognized since the 1940s, and inositol is known to have important metabolic roles, such as in fat metabolism and cell function. Inositol is also combined with phosphate at a cellular level to recreate phytate (which acts as a potent antioxidant in the cell) and the lower phytate esters (IP3 and IP4 are important to cell function).
In fact, the latest research has shown that inositol promotes growth and feed efficiency regardless of whether the diet is deficient in P. The exact reason is not known, but a recent broiler trial involving the addition of inositol to the diet at levels equivalent to all dietary phytate being degraded has confirmed its important role in the overall superdosing response.
Superdosing challenge
The challenge when implementing a superdosing strategy is therefore to achieve as near complete degradation of phytate to inositol as possible, a task that is considerably harder than the partial breakdown needed to release the usual 0.15 percent available P (AvP) target of standard phytase dosing. As such, superdosing places an even greater emphasis on phytase efficiency and the characteristics necessary to drive phytate to near elimination and so maximize inositol release.
Thermostability to withstand high temperature feed processing and protect activity is essential, ideally without the need for a coating that could slow the onset of activity in the stomach. This is where phytate is soluble, and so available for degradation, under acidic conditions.
Good gastric tolerance and stability is also needed to resist breakdown by the bird’s own digestive enzymes, while phytase activity should be optimized at the low pH of the stomach (pH 2-3). Further, high rate and extent of phytate breakdown are essential – fast enough to prevent anti-nutrient effects taking place and continuing even at low concentrations to drive toward complete elimination.
Additional P release
The logical question raised by this level of phytate destruction is that of potentially increased matrix values in order to benefit – through further diet cost reductions – from the additional P released compared to standard phytase dosing.
However, experience to date has shown that it is not practically possible to guarantee the release of 0.20-0.25 percent AvP, for example, when feed ingredients (and thus diet phytate-P concentrations) vary. The P requirements of superdosed birds also may be higher due to factors such as the subsequent increase in growth rate and the use of P to re-phosphorylate the additional available inositol at a cellular level.
Optimised phytase
Practical experience during the past two or so years has shown that the overall financial and performance benefits that come from phytase superdosing are only maximized when a standard mineral matrix is used. The focus should instead be on choosing a phytase capable of efficient and extensive phytate destruction to eliminate anti-nutrient effects and release inositol.
This in itself is not straightforward, because the range of commercial phytase products available has grown considerably in recent years. Successful phytase superdosing therefore depends as much on making the right choices and buying decisions as it does on correct dosage and application.
A better understanding of the factors and mechanisms that underpin the response in the bird, and the emerging role of inositol in delivering that response, therefore represents a key step in the journey toward achieving consistent superdosing results.

















